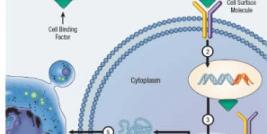
Molecular Biology of the Retrovirus
Retroviral particles typically comprise two identical copies of single-stranded, positive sense RNA as a genome, ranging between 7 – 10 kb, which is contained within a protein shell (capsid). The capsid is surrounded by a lipid membrane from which extrudes envelope glycoproteins resulting in a total particle size between 90 and 140 mm. It is the envelope proteins that confer retroviral specificity, as they are responsible for attachment to the target cell allowing for virus internalisation. The envelope-dependent tropism of retroviruses is commonly categorised into three groups; ecotropic (infects murine cells only), xenotropic (infects all cell types except those of murine origin) and amphotropic (infects cells of both murine and non-murine lineage).
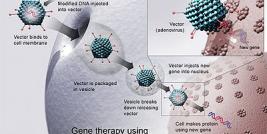
Subsequent to viral entry into the cell, the RNA genome is reverse transcribed into double-stranded cDNA by the retroviral reverse transcriptase. Reverse transcribed cDNA then translocates to the nucleus and integrates into the host cell genome through the action of retroviral integrase. Therefore, retroviral-mediated transduction (the term used to describe retrovirus infection) of target cells results in stable integration of the viral genome. This is perhaps the most attractive feature of the retrovirus as a gene transfer vehicle for gene therapy, due to the requirement of permanent therapeutic transgene expression.
During the life cycle of wild-type retrovirus, production of new particles arises subsequent to the transcription of integrated viral DNA (provirus). The core proteins assemble in the cytoplasm at the plasma membrane where the RNA genome is targeted to the capsid by the packaging signal (psi). The retroviral genome is divided into three regions: gag, pol and env encoding for capsid proteins, viral enzymes and envelope proteins, respectively (figure 1A). The structural genes encoded by the gag gene (group antigen gene) are expressed in the form of a polyprotein yielding four proteins: p10 (nucleocapsid), p12, p15 (matrix protein) and p30 (capsid protein). The polymerase (pol) gene, lying downstream of the gag gene, encodes the integrase, reverse transcriptase and the protease enzymes. Finally, the envelope (env) gene, which is downstream of the pol gene, also encodes a polyprotein that is cleaved by viral protease to yield gp70 (surface glycoprotein) and p15E (transmembrane protein). The entire retroviral genome encoding these structural and enzymatic proteins is flanked by two long terminal repeats (LTRs) (see diagram one).
Diagram One: Retrovirus Genome
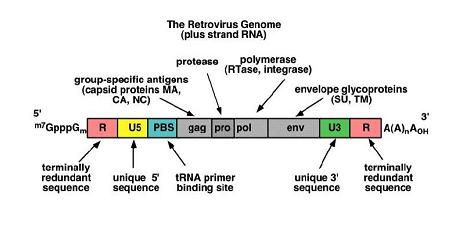
The LTRs are essential for the initiation of viral DNA syntheses, integration of proviral DNA and the regulation of viral gene expression. Finally, the psi located just upstream of the gag region is an essential cis element during retroviral particle production. The most commonly utilised retrovirus for gene transfer applications is that based on the Moloney Murine Leukaemia Virus (MoMLV), which has perhaps the simplest genome of all the retroviruses, thus making it ideal for modification and use as a gene transfer agent.
Recombinant Retroviral Vectors
First Generation
Retroviruses are made replication incompetent and suitable for gene transfer applications by the removal of the gag, pol and env genes, which are subsequently replaced by an expression cassette containing the transgene of interest (see diagram two).
Diagram Two: Retroviral Vector
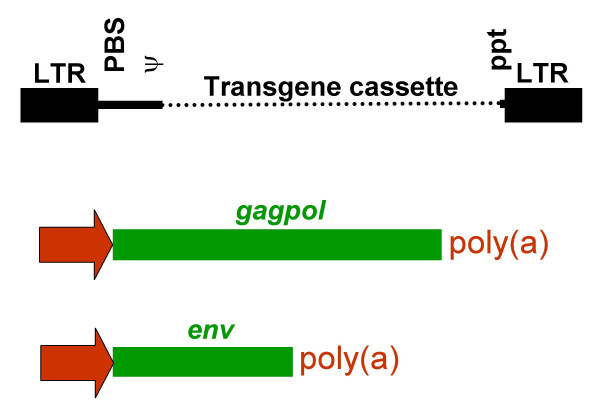
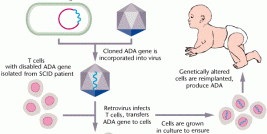
The essential LTR and psi elements are retained in order to allow transgene expression and packaging into viral capsids during vector production. As a result retroviral vectors are still capable of transducing a cell and expressing the foreign gene, but further viral particles can no longer be produced due to the lack of the structural and enzymatic genes. In order to facilitate vector production, the gag, pol and env genes are introduced into murine or human cell lines to make retroviral-packaging or -producing cells.
These cell lines are then transiently or stably transfected with plasmid DNA encoding the retroviral vector genome containing the therapeutic transgene. Subsequent to transfection, the viral genome is expressed, packaged and released by the cell as a replication-incompetent retrovirus (see diagram). Retroviral vector is then harvested from the culture medium and purified by ultracentrifugation generating vector of titres in the range of 106 to 107 cfu/ml (colony forming units; as determined by the ability of vector to stably transduce target cells generating vector-containing cell colonies). This is quite a low yield when compared to the production of vectors based on other virus types. However, there are extensive efforts to increase vector production from retroviral-producer cell lines by altering the temperature and pH at which producer cells are cultured, or by the addition of compounds that increase expression from the retroviral LTRs e.g. sodim butyrate or by the use of advanced cell culture facilities, e.g. the cell cube, microcarrier suspension culture system and the packed or fluidised bed.
Second Generation
Production of retroviral vector from stable cell lines is associated with a risk of the generation of replication competent retrovirus (RCR) that arises subsequent to the recombination of the different retroviral elements in the producer cells. To avoid this phenomenon the viral elements are split onto at least three separate plasmids, so that one plasmid contains the therapeutic transgene-expressing viral genome, a second expresses the gagpol polypeptide and the third expresses the envelope protein. By doing this three separate recombination events are required to generate RCR, something that is highly unlikely. Despite the separation of these elements onto separate plasmids there will always be some risk of homologous recombination. This is because the pol and env genes are out of frame and share some common sequences, as do the psi and gag gene. In order to avoid this potential recombination more recent generations of retroviral vectors contain envelope proteins derived from retroviruses isolated from a different species with dissimilar env sequences. The result of these modifications is the generation of a vector capable of highly efficient and stable transgene expression perfect for the gene therapy of many dominantly inherited monogenic disorders.
However, retroviral vectors do have some limitations. Firstly, they can only transduce rapidly dividing cells, as the reverse transcribed retroviral genome cannot enter the cell’s nucleus unless the nuclear membrane is disrupted during mitosis. Secondly, subsequent to integration, the retroviral LTR that drives transgene expression is often silenced by promoter methylation thus gene expression is transient in these cases. However, the most severe limitation of the use of retroviral vectors is the fact that they integrate into the host genome in a random manner. This raises the potential of insertional mutagenesis or the activation of certain oncogenes that may be adjacent to the strong retroviral promoter. Most of these problems can be overcome by the elegant manipulation of the viral genome by molecular biological techniques. For instance, it is possible to create self-inactivating retroviral vectors in order to reduce the potential for tumourigenesis following random integration. In this approach, a section of the 3’ LTR of the virus is deleted so that it cannot drive expression of genes close by the integration site. A weaker tissue specific promoter can then be introduced to drive the expression of the therapeutic gene. However, not until some degree of site-specific integration is afforded to retroviral vectors, will they be considered as the ideal gene therapy tool. In spite of this final limitation, retroviral vectors remain one of the most commonly used viruses in gene therapy clinical trials.